Abstract
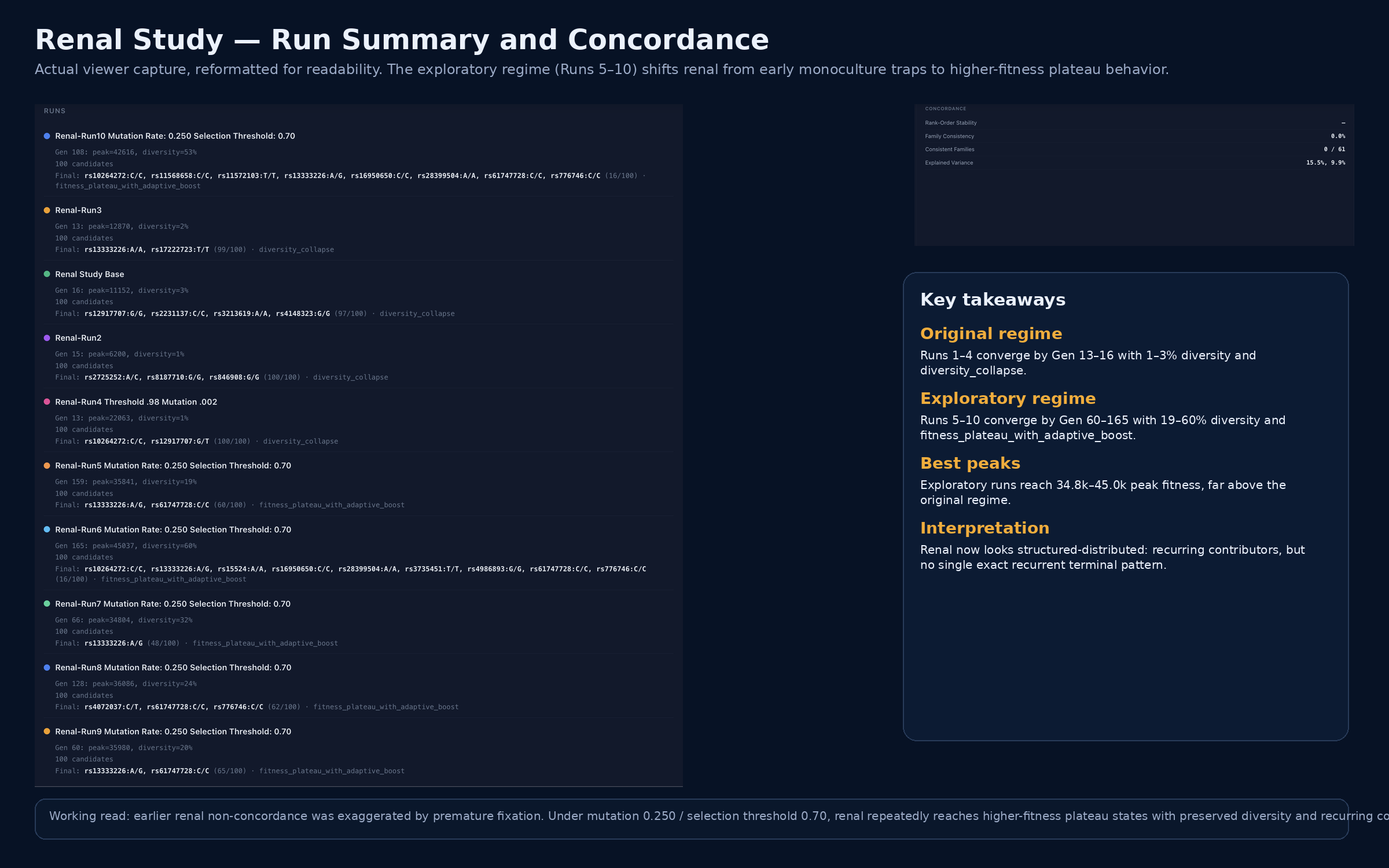
Renal phenotype simulations conducted under the original search regime converged prematurely into low-diversity monocultures, obscuring cross-run structure and exaggerating apparent instability. To determine whether these outcomes reflected true biological architecture or search-regime artifact, mutation rate was increased and selection pressure was reduced. Under these exploratory conditions, the system shifted into a distinct behavioral regime characterized by later convergence, substantially higher peak fitness, preserved diversity, and termination by fitness plateau with adaptive boost rather than immediate diversity collapse. Although exact terminal configurations remained variable across runs, recurrent contributors emerged repeatedly, including rs13333226, rs61747728, rs776746, rs10264272, rs16950650, and rs28399504. These findings indicate that renal is neither reducible to a single dominant configuration nor random, but instead behaves as a structured distributed phenotype under exploratory conditions.
Introduction
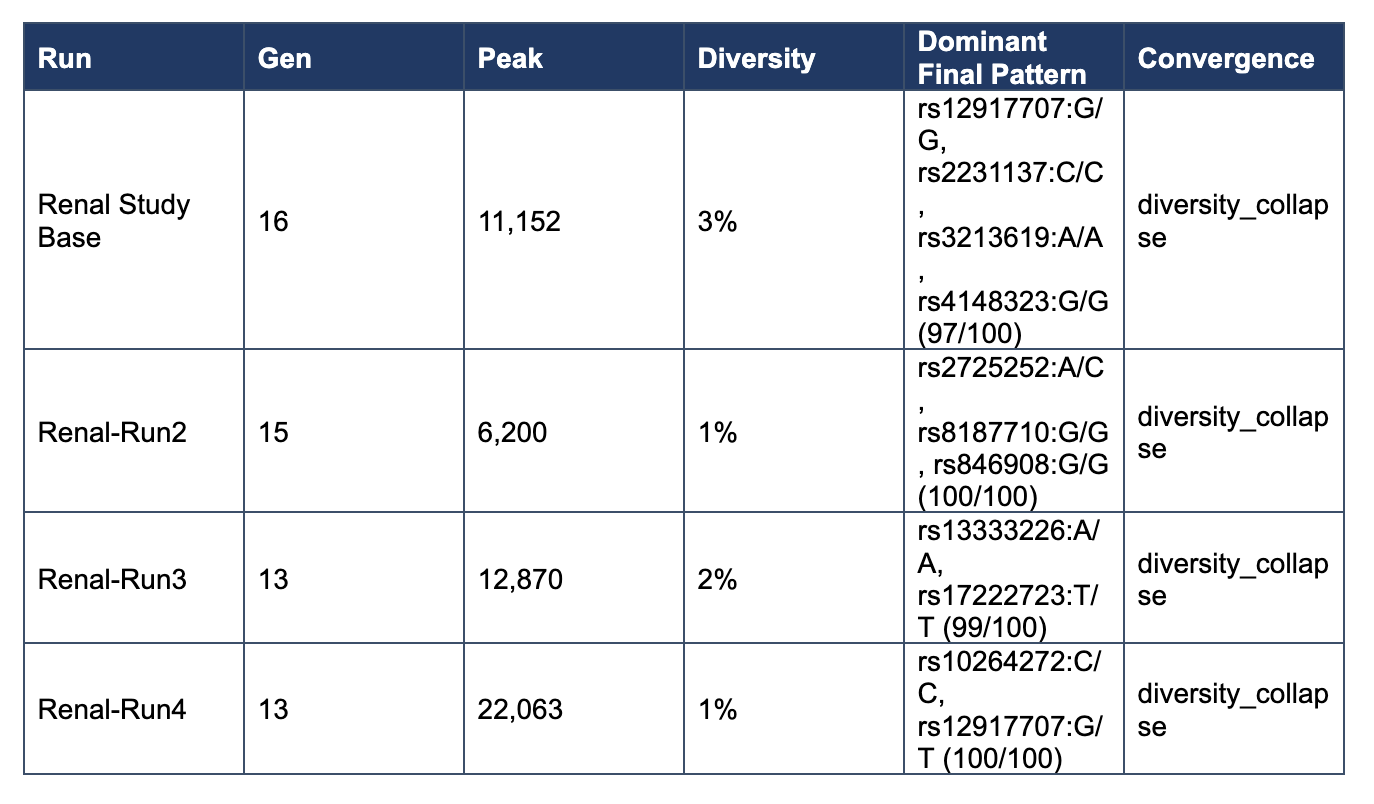
Initial renal simulations converged rapidly into low-diversity terminal states, often by generations 13–16, with different attractors emerging across independent runs. These outcomes raised a central question: did they reflect genuine biological multi-stability, or were they artifacts of an overly restrictive search regime?
To address this, a more exploratory parameterization was tested to preserve search diversity and reduce premature fixation. The objective was to determine whether renal would remain unstable, collapse into one dominant recurrent configuration, or reveal a more distributed but biologically coherent architecture.
Methods
Original Regime
Baseline mutation and selection settings
Typical convergence by generations 13–16
Terminal diversity of 1–3%
Convergence reason dominated by diversity_collapse
Exploratory Regime
Mutation rate: 0.250
Selection threshold: 0.70
Later convergence with preserved diversity
Convergence reason dominated by fitness_plateau_with_adaptive_boost
For each run, the following metrics were recorded: generation at convergence, peak fitness, diversity at termination, dominant final pattern and dominant count, and recurrence of contributors across runs.
Results
Regime A: Premature Fixation
Under the original search settings, renal runs converged by generations 13–16 with terminal diversity of 1–3%. Final states were near-monocultures, with 97–100 of 100 candidates occupying a single dominant pattern. However, the dominant attractor differed across runs, indicating rapid fixation without cross-run structural recurrence.
Regime B: Exploratory Plateau
Under the exploratory regime, renal simulations shifted into a qualitatively different behavioral class. Runs converged later, reached substantially higher peak fitness, preserved much greater diversity, and terminated by fitness plateau with adaptive boost rather than immediate diversity collapse.
Table 1. Exploratory Regime Summary
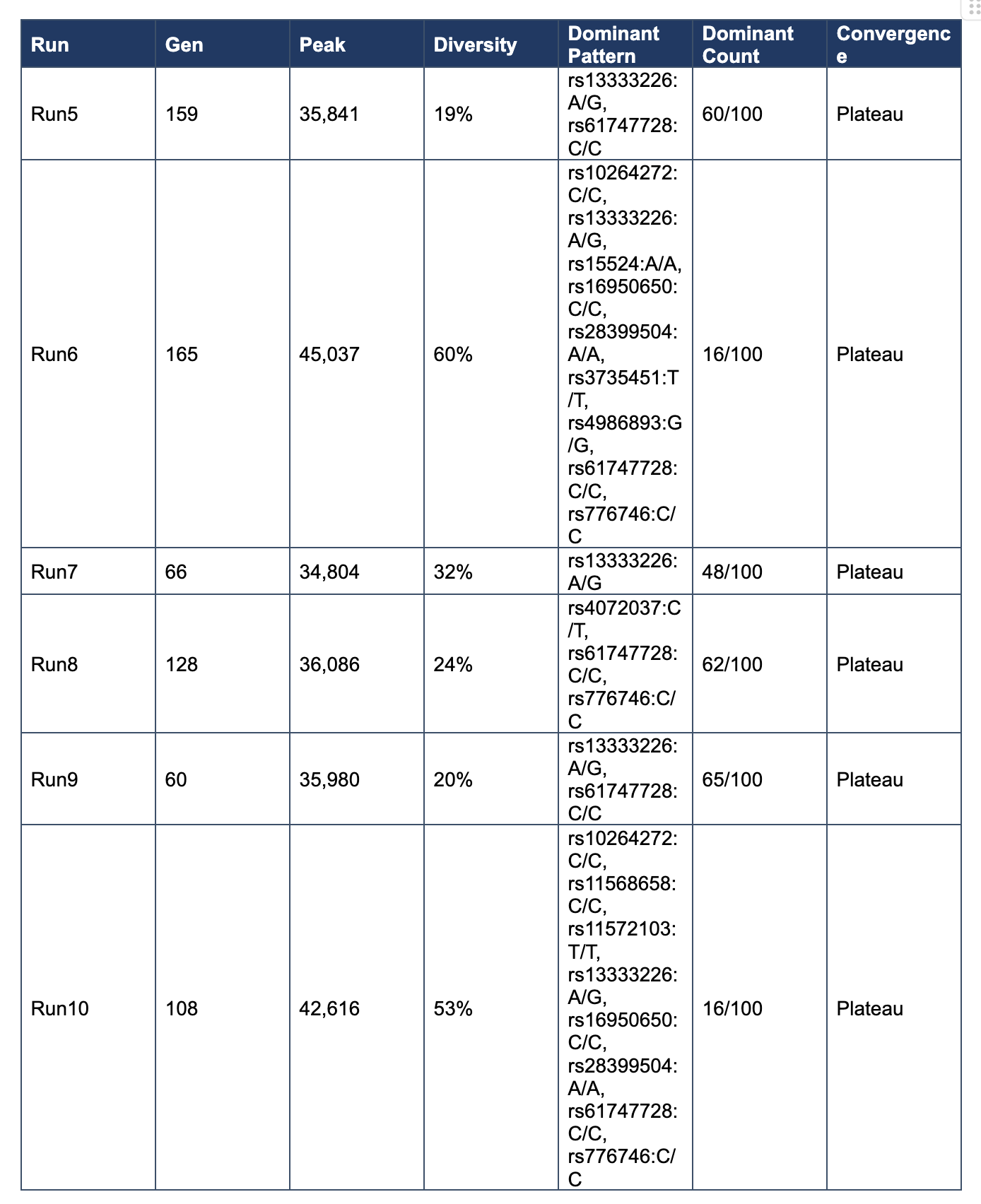
Across Runs 5–10, convergence occurred between generations 60 and 165, diversity was preserved at 19–60%, and peak fitness increased to a range of 34,804–45,037. Exact terminal patterns remained variable, but recurrent contributors emerged repeatedly across runs.
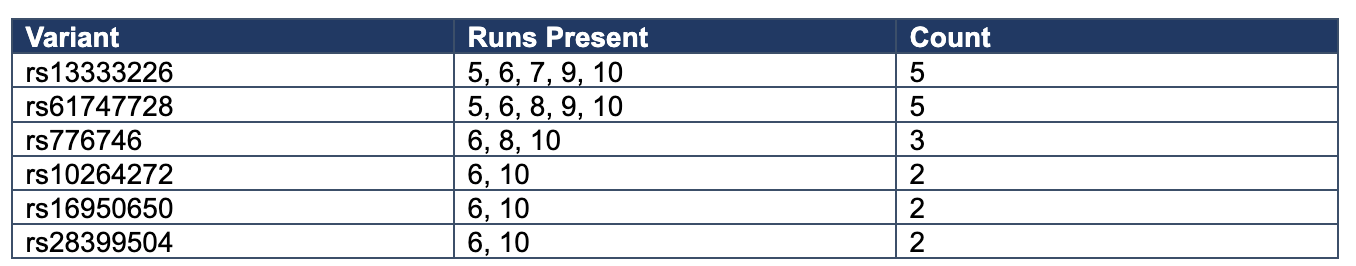
Table 2. Recurrent Contributors Across Exploratory Runs
Discussion
These findings indicate that renal is not reducible to a single dominant recurrent configuration under the tested conditions. At the same time, the exploratory runs do not behave randomly. Instead, they repeatedly recover related high-fitness configurations with overlapping contributors, indicating a structured distributed architecture. The sharp contrast between the original and exploratory regimes further suggests that the earlier low-diversity renal outcomes were at least partly artifacts of premature fixation. Under healthier exploration, renal reveals coherent contributor families rather than exact-pattern recurrence.

Figure 1. Renal simulations exhibit two distinct behavioral regimes.
Caption:
Renal domain separates into premature-fixation and exploratory distributed regimes. Under the original renal simulation settings, independent runs collapsed by generations 13–16 into low-diversity monocultures via diversity_collapse, with different dominant attractors across runs. Under the exploratory regime (mutation 0.250, selection threshold 0.70), runs converged later, reached substantially higher peak fitness, preserved markedly greater structural diversity, and terminated by fitness_plateau_with_adaptive_boost rather than immediate collapse. Exact terminal patterns remained variable, but recurrent contributors repeatedly emerged across exploratory runs, indicating a structured distributed architecture rather than random instability. PROTEUS explicitly defines such outcomes as computationally measurable properties of simulation output under defined conditions, distinguishing reducible from distributed systems by concordance, structural heterogeneity, and recurrence behavior.
Under the original search settings, renal runs converged by generations 13–16 through immediate diversity collapse, producing low-diversity monocultures and different terminal basins across runs. Under the exploratory regime (mutation 0.250, selection threshold 0.70), runs converged substantially later, retained markedly higher diversity, and terminated by fitness plateau with adaptive mutation rather than early collapse. This indicates that the earlier renal result was at least partly a search-regime artifact and that renal behaves differently under healthier exploration conditions. PROTEUS explicitly treats convergence behavior as a measured property of the simulation output, distinguishing reducible from distributed systems rather than assuming either class a priori.
Cross-run overlay of renal exploratory runs. Shared embedding of independent renal simulations under exploratory settings shows that exact terminal configurations differ across runs, but candidate populations repeatedly occupy related high-fitness regions associated with recurrent contributors including rs13333226, rs61747728, and rs776746. This pattern supports family-level recurrence without exact-pattern recurrence, consistent with the distributed configuration class described in PROTEUS.
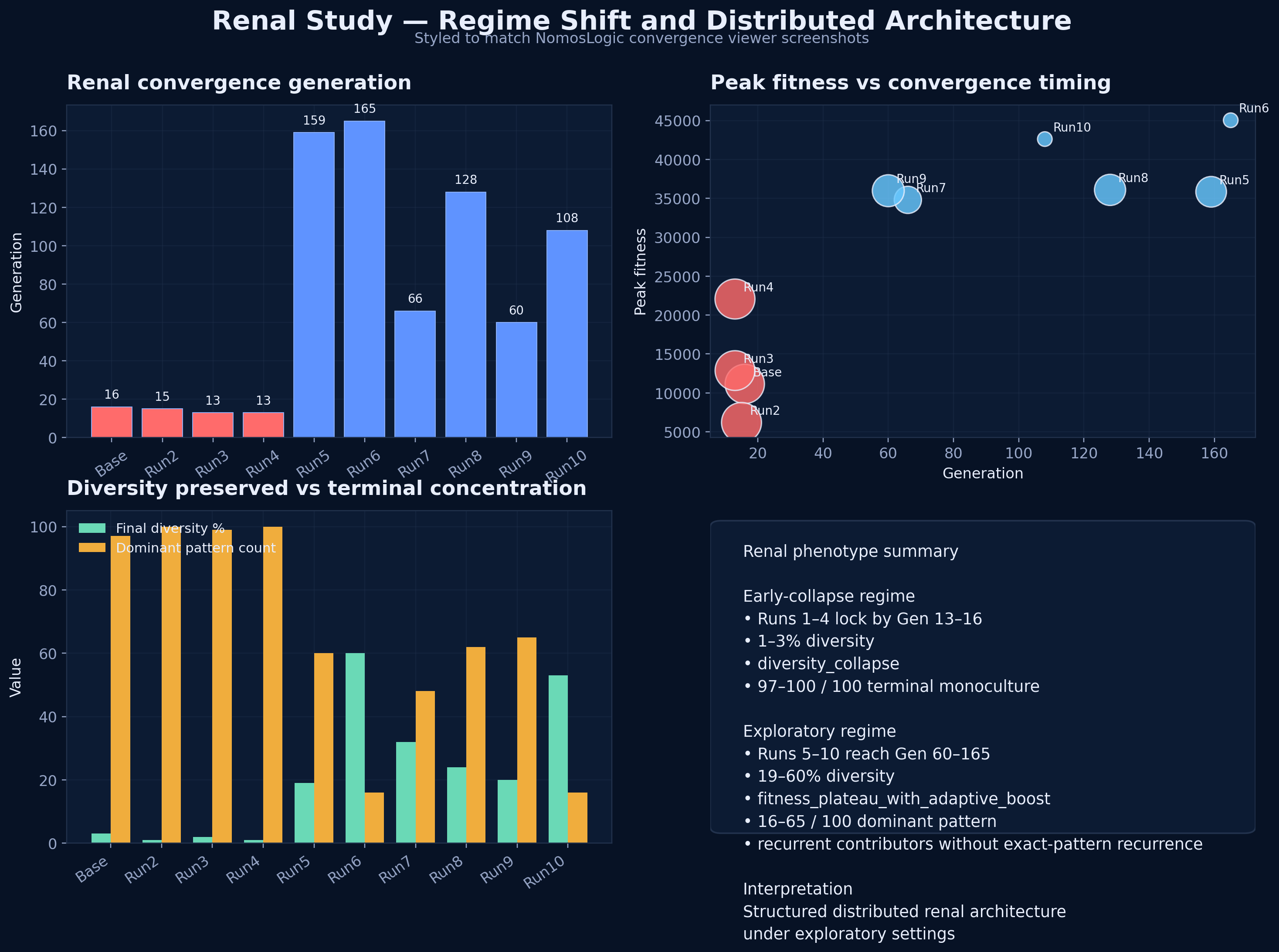
Figure 2. Exploratory renal runs achieve higher fitness while preserving substantially more diversity.
Caption:
The original renal settings produced early low-fitness monoculture collapse, whereas the exploratory settings yielded later plateau behavior, much higher peak fitness, and sustained structural diversity. This shift argues against interpreting the early renal runs as the definitive phenotype and instead supports a parameter-sensitive distributed architecture under more adequate exploration.
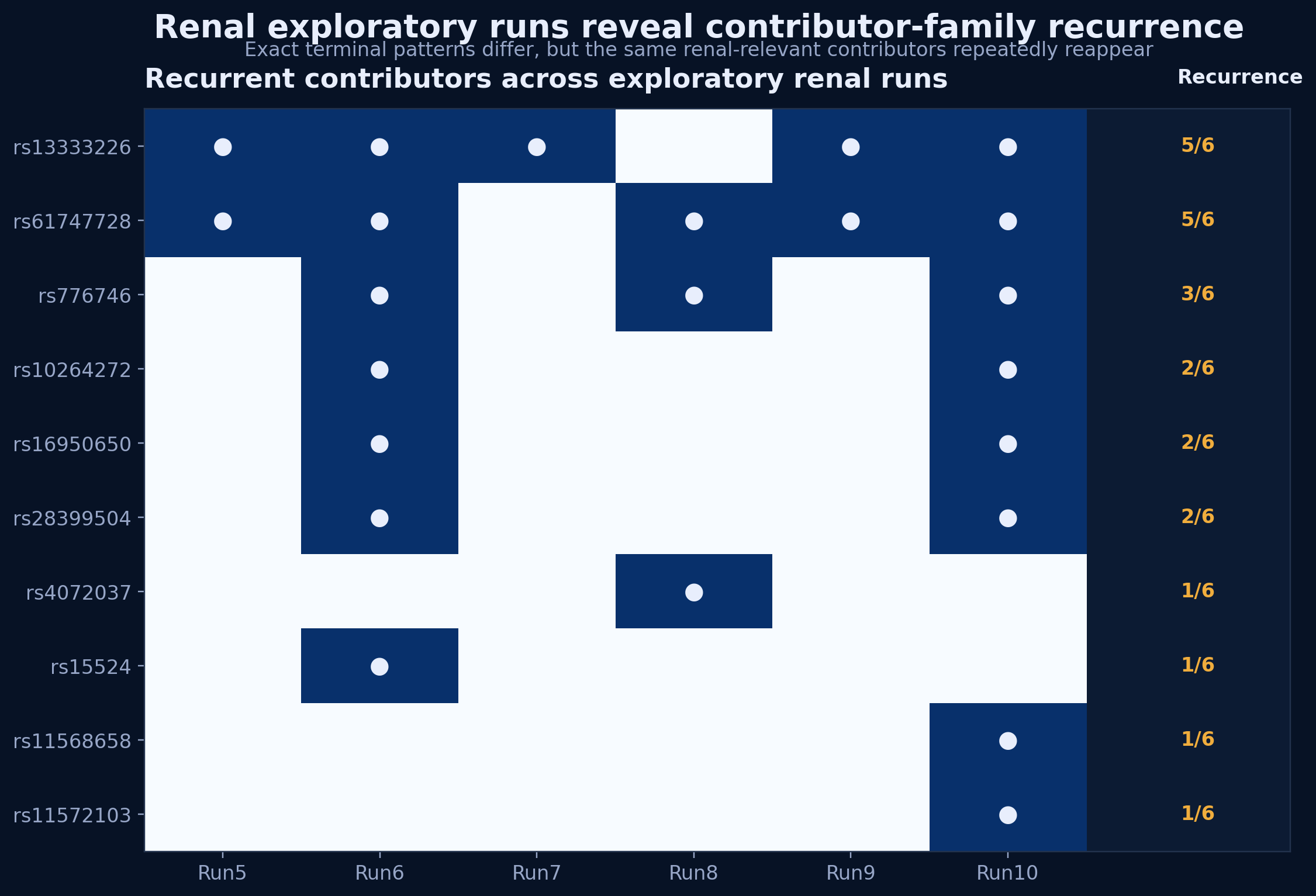
Figure 3. Exploratory renal runs do not recur to one exact terminal pattern, but repeatedly recover a shared contributor family.
Caption:
Across Runs 5–10, exact dominant configurations remain variable, but recurrent contributors emerge repeatedly, especially rs13333226 and rs61747728, with secondary recurrence of rs776746, rs10264272, rs16950650, and rs28399504. This pattern supports family-level recurrence and biological coherence without exact-pattern recurrence, consistent with a structured distributed architecture rather than a reducible single-attractor system.
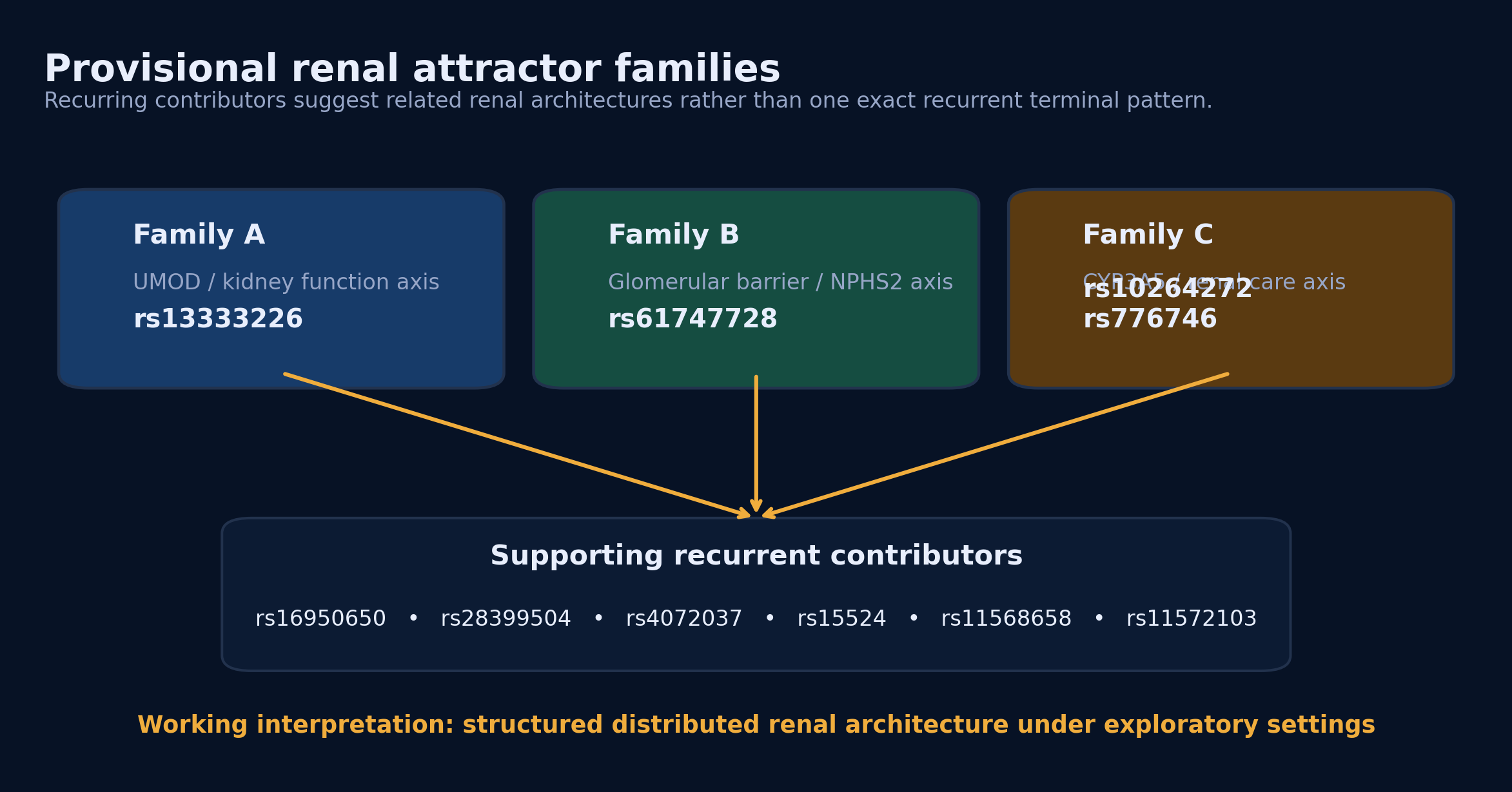
Figure 4. Renal exploratory runs resolve into related attractor families rather than one exact recurrent terminal configuration.
Caption:
Recurrence analysis suggests at least three renal-relevant contributor axes centered on UMOD-associated, glomerular barrier-associated, and CYP3A5-associated biology. Different runs assemble distinct high-fitness combinations from this shared contributor neighborhood, indicating a distributed but biologically coherent renal architecture. This is consistent with the PROTEUS distinction between reducible systems, which recur to one dominant configuration, and distributed systems, which preserve stable behavior across multiple related high-fitness configurations.
Conclusion
Renal behaves as a structured distributed phenotype under exploratory search conditions. Earlier interpretations based on rapid monoculture collapse were likely distorted by premature fixation. When exploration is preserved, renal consistently reaches higher-fitness plateau states with sustained diversity and recurring contributors, supporting a distributed but biologically coherent system architecture.
Appendix: