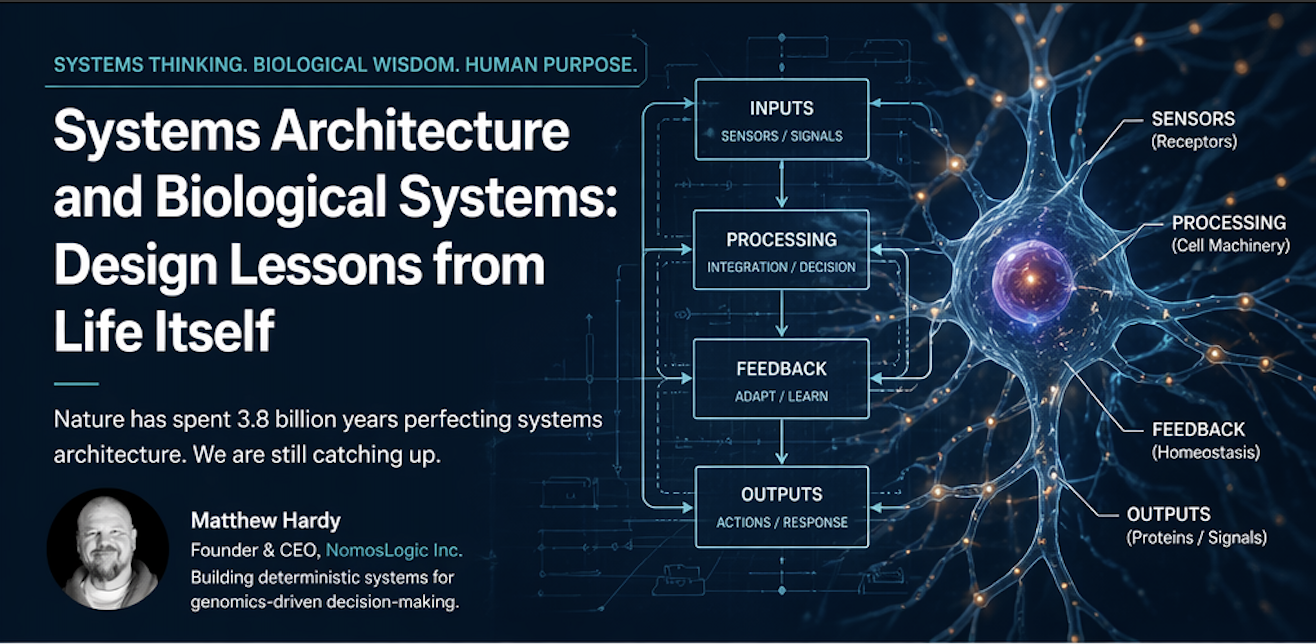

There is a strange arrogance in modern systems design.
We build software, infrastructure, networks, and operating models as if the hardest architecture problems were invented in the last fifty years. We talk about resilience, fault tolerance, observability, feedback loops, graceful degradation, and adaptive control as if these are new ideas. They are not new. They are ancient. Biology solved them first.
Long before we wrote the first line of code, life was already engineering complex, distributed, self-correcting systems under impossible constraints. It had to operate without central coordination, survive partial failure, respond to uncertain inputs, preserve continuity through change, and make decisions in real time with incomplete information. In other words, biology has been solving the exact class of problems that systems architects struggle with today.
That matters more than most people realize.
Because if we want to build durable systems, we should study the one architecture that has survived every stress test imaginable: life itself.
The central lesson is simple. Biological systems are not collections of isolated parts. They are networks of constrained relationships. A gene does not act alone. A cell does not function alone. An organ does not survive alone. A human being does not exist apart from the environment that shapes and pressures the system. The same is true in systems architecture. Services, databases, pipelines, interfaces, and workflows are not meaningful merely as components. They become meaningful through their interdependence.
That is the first design error many organizations make. They optimize the node and ignore the network.
A team improves a model. Another rewrites a service. Another hardens a database. Another adds automation to a deployment pipeline. Each local improvement may be rational. Yet the larger system becomes more fragile because the architecture of interaction was never addressed. Local optimization without systemic understanding creates hidden brittleness. Biology rarely makes that mistake. It does not optimize organs in abstraction. It stabilizes relationships across the whole.
This is why biological systems should force us to think differently about architecture. The question is not only whether a component is good. The question is whether the system can absorb variation, error, mutation, and environmental change without losing coherence.
That is what resilience actually is.
In engineered systems, resilience is too often treated as a feature. In biology, resilience is structural. It emerges from redundancy, layered signaling, feedback, buffering, modularity, and adaptation. A biological system does not survive because it is perfect. It survives because it is designed to continue functioning when perfection fails.
That is the second lesson. Redundancy is not waste. It is survival logic.
In business environments obsessed with efficiency, redundancy often looks indulgent. Duplicate pathways, fallback mechanisms, extra signals, overlapping controls, and alternative routes are seen as cost centers. But biology tells a different story. Redundancy is what keeps a system alive when one path fails. It is what allows partial damage without total collapse. It is what creates reserve capacity.
In software and platform design, the same principle holds. If a single service failure can bring down the whole environment, the system is not elegant. It is underdesigned. If a single executive departure, single vendor dependency, single data source, or single decision layer can destabilize the enterprise, the architecture is not strong. It is brittle.
Biological systems build reserve into the design because they assume disruption is normal. Good systems architects should do the same.
The third lesson is feedback.
Life is governed by continuous sensing and response. Biological systems do not wait for annual review cycles. They measure, adjust, compensate, suppress, amplify, and reorient constantly. They are not static machines. They are adaptive systems.
This should sound familiar, because the best modern architecture patterns are converging on the same truth. Systems that cannot observe themselves cannot correct themselves. Systems that cannot correct themselves cannot remain stable. Systems that cannot remain stable eventually fail.
But there is a deeper layer here. Feedback is not merely informational. It is formative. It changes the future state of the system. In biology, feedback is the mechanism by which order persists under stress. In engineered systems, feedback should play the same role. Metrics should not exist for dashboards alone. Signals should not exist merely for reporting. They should exist to alter behavior.
That is what distinguishes observability from architecture. A dashboard that informs no response is decorative. A signal that changes the system is architectural.
The fourth lesson is that mechanism matters more than surface output.
A great deal of modern decision-making, especially in technical and commercial systems, is driven by confidence language. Probabilities, scores, rankings, predictions, likelihoods. These are useful, but they are incomplete. Confidence may tell you what to believe. It rarely tells you what to do.
Biology operates differently. It is built on mechanism. Signals flow through pathways. Inputs alter expression. Structure shapes consequence. Function emerges from causal interaction, not just statistical association.
That has profound implications for both technology and medicine. A system that can only say something is likely important is inherently weaker than one that can explain why it matters, how it propagates, and what happens if it fails. Mechanism is actionable. Mechanism can be audited. Mechanism can be taught. Mechanism can be defended.
This is why the future of serious systems architecture, especially in biology and medicine, cannot stop at black-box prediction. It has to move toward interpretable structure and governed mechanism.
The fifth lesson is adaptation.
One of the great failures of static architecture is the assumption that the environment is stable enough for fixed design. Biology never makes that assumption. Biological systems are shaped by changing pressures. They adapt because the world changes. Selection, compensation, and reconfiguration are not edge cases. They are the normal condition of life.
Most engineered systems still behave as if architecture is complete once it is deployed. But real systems are never finished. They are always under pressure from scale, competition, regulation, cost, misuse, drift, mutation, and time. The architecture that survives is not the one that predicts every future state. It is the one that can reorganize without losing identity.
That is a critical distinction.
A fragile system interprets perturbation as failure.
A durable system interprets perturbation as information.
Biology does the latter. That is why it persists.
The sixth lesson is constraint.
Engineers often think freedom creates better systems. Biology suggests the opposite. Constraint is what gives systems form. Cells function because their behavior is bounded. Organisms survive because signaling, energy, repair, and replication occur inside governed limits. Remove the constraints and the system does not become more powerful. It becomes chaotic.
The same is true in architecture. Governance, interfaces, protocols, sequencing, validation, and bounded responsibility are not bureaucratic overhead when done correctly. They are the skeleton that keeps complexity from dissolving into noise. Constraint is not the enemy of innovation. It is the condition that makes reliable innovation possible.
This matters at the organizational level too. Companies often confuse speed with velocity. Speed is motion. Velocity is directed motion. Biology favors the second. Systems architecture should as well.
The final lesson may be the most important. Beauty in systems is not decoration. It is coherence.
A beautiful biological system is not beautiful because it is visually pleasing. It is beautiful because its structure is truthful. Form, function, adaptation, and continuity align. The same is true in architecture. The most elegant systems are not the ones with the most impressive diagrams or the most fashionable abstractions. They are the ones where the design reflects reality, where the constraints are honest, where the interfaces make sense, and where the system holds under pressure.
That is the standard.
If we want to build infrastructure that lasts, whether in software, enterprises, or medicine, we should stop pretending architecture begins with us. Life has already written the deeper playbook. Distributed control. Redundancy with purpose. Mechanism over opacity. Feedback that changes behavior. Adaptation under pressure. Constraint as structure. Coherence as beauty.
The work now is not to invent these principles.
It is to learn them well enough to build in accordance with them.
Because the best systems are not merely efficient.
They are alive to reality.